
5.4: Lipid Probes
- Page ID
- 17563
Lipids are characteristically different from proteins, making in vivo identification and labeling more difficult. Compared to protons that are character by unique sequences of amino acids and that are often interfaced with hydrophilic regions, lipids are less recognizable due to their relatively inconspicuous hydrocarbon structures and their preference for hydrophobic environments. Additionally, the complexity of lipid biosynthesis prevents the use of techniques analogous to simplifying protein research, such as overexpression and radiolabeling, as many different downstream lipid species may be affected due to reliance on the original biosynthesis pathway. These major differences contribute to the difficulty of finding and using lipid probes. One of the first notable techniques for tagging lipids originated in the early 1980’s when several experiments involving one of the fatty acids in phospholipid and glycosphingolipids-like analogues, which were replaced by 6 carbon tethers that were also connected to a fluorescent probe (see research associated with Klausner and Karnovsky or Stierr and Stackmann [1,2,3,4]). Lipid labeling offers an alternative to destructive methods that use detergents, such as the formation of homogeneous vesicles that are characteristically different than the native environments for most lipids, to induce phase separation or separate membranes, allowing for measurement made in native environments without immobilization [5]. There are three major types of lipid probes: fluorescent probes can be used for identifying target membranes or lipids [1,2,3,4], spin-probes are useful for measuring membrane dynamics [27], and isotopic probes are often used for membrane composition analysis [33].
Fluorescent Lipid Probes
Fluorescent probes are molecules associated with lipid systems that can be excited to emit energy that can be measured to identify and track specific lipids. They can be divided into three categories: probes that target lipid components (such as gangliosides), probes that flexibly associate with lipids despite their phase, and probes that have ordered or disordered phase preferentiality [5]. Commonly used fluorescent probes are shown below in Table 1, where the spectroscopic properties and phase preference are included, and Figure \(\PageIndex{1}\) above demonstrates the visual response of fluorescent probes.

The first of these more specific groups consists mainly of fluorescent proteins that target lipid components, like cholesterol, through inactivation mechanisms; this is the least populated group of lipid probes [6,7]. These offer bright and photostable characteristics that are not very sensitive to environmental polarity or viscosity and are often the optimum dye to use when working with fluorescence correlation spectroscopy, single molecule imaging and other super-resolution techniques [8,9,10,11].
The second group includes lipid-like dyes that associate with lipid membranes due to labeling of lipids or lipophilic structures of non-lipid molecules that can demonstrate similar phases of biological membranes, such as the ordered and disordered phases seen with lipid rafts [12]. Fluorescent lipid probes are chemical markers that can bind to certain functional groups that belong to one of the two amphiphilic structures found in lipids: the hydrophilic headgroup or the hydrophobic tail. The function of study for the lipid will depend on the choice of dye as the additional probe will chemically affect the local functionalization by changing hydrophilic/phobic balance while also sterically impeding lipid packing in membranes [13]. Head group-associated probes are useful for research where lipid head groups play a significant role, such as lipid sorting, while chain-associated probes are useful for cases when chain regions are important, like studying membrane fluidity [14, 15]. The insertion of these marked lipids into the membrane or system of study is then facilitated by the lipid base of the probe. These tagged lipids have preferential association with certain membrane phases and thus will show varying fluorescence depending on the environment, as well as affecting membrane stability [16, 17, 18]. Additionally, lipophilic probes that consist of non-lipid structures can take the form of long-chain hydrocarbons (LCH dyes) or poly-cyclic aromatic hydrocarbons (PAH dyes) can also be used [5]. LCH dyes include alkylated cyanines and rhodamines, which have more similar structures to lipids than PAH dyes that consist mainly of neutral aromatic compounds that lack alkyl chains [5]. These probes are often used with fluorescence correlation spectroscopy and single molecule imaging [19, 20].
The third group of dyes, commonly referred to as environment-sensitive dyes, respond spectroscopically to local environment properties like polarity, hydration, viscosity and pH while also being able to distinguish between ordered and disordered membrane phases [21, 22]. These can be further categorized into solvatochromic probes that have fluorescence that responds to the environmental polarity and viscosity-sensitive, molecular rotor probes that vary their fluorescence intensity and duration with local viscosity. Solvatochromic dyes rely on changes in dipole-dipole or hydrogen bonding interactions with their environment that affect the maxima of their excitation/emission spectra to give optical responses. These probes are used most often with ratiometric or lifetime measurements due to the change in their emission spectra and lifetime as a response to their local membrane phase [15, 23]. Molecular rotors rely on local lipid interactions, which are directly related to membrane viscosity to affect the intramolecular rotation of the fluorophore and thus the fluorescence quantum yield (or the ratio of number of photons emitted to the number of photons absorbed) [24, 16]. These probes are also used with fluorescence lifetime measurements due to the relationship between their fluorescence lifetime and local viscosity [25, 26].
Spin Probes
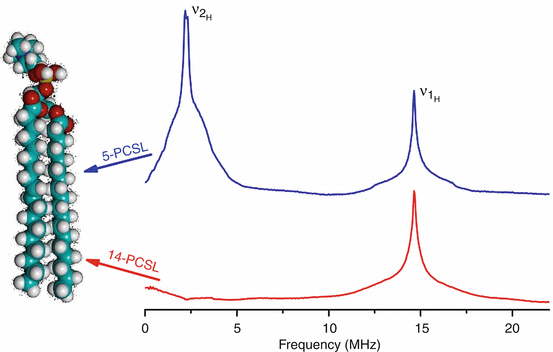
A spin probe is molecule with an unpaired electron that can associated to another molecule and can be used with electron paramagnetic resonance (EPR) spectroscopy to identify lipid membrane dynamics. Common species include nitroxide radicals associated with either the hydrophilic headgroup or the hydrophobic chains of the lipid molecules that can then be identified by EPR to determine membrane fluidity through correlation with line-width and line-shape, which can describe local environmental influences on the labeled lipids, as shown in Figure \(\PageIndex{2}\) [27]. Other available spin probes applicable to lipids include TEMPO ((2,2,6,6-tetramethylpiperidin-1-yl)oxyl) and DTBN (di-tert-butyl nitroxide), which are notably water-soluble and can interact with the aqueous-hydrocarbon interface [28]. The main targeting mechanism for introducing these spin probes to native environments involves adding spin labeled precursors that will be used in lipid biosynthesis pathways, which will then be integrated in target membranes [29]. Spin labels are used in low concentrations in lipid systems to avoid spin-spin interactions and prevent membrane disruption. Common experiments involve measuring the response of chain-labeled lipids as the labeling group moves down the acyl chain of lipid segments away from the headgroup, which demonstrates an increase in spin the further away with increasing distance from the headgroup [30, 31, 32].
| Name | Absorbance Max (nm) | Fluorescence Max (nm) | Partitioning in giant unicellular vesicles | Partitioning in giant plasma membrane vesicles |
|---|---|---|---|---|
| Cholesterol derivatives | ||||
| TF-Chol | 495 | 507 | Lo (A, 80%) | Lo (66%) |
| NBD-Chol | 470 | 538 | Ld (A) | – |
| Cholestatrienol | 324 | 390 | Lo (A) | – |
| PE head group labeled | ||||
| NBD-DOPE | 470 | 538 | Ld (A) | Ldb |
| NBD-DPPE | 470 | 538 | Lo/Ld (A-C) | Lo |
| Rh-DOPE | 560 | 583 | Ld (A) | Ldb |
| Rh-DPPE | 560 | 583 | Ld (A) | Ldb |
| Texas Red-DPPE | 595 | 614 | Ld (A) | – |
| DSPE-KK114 | 640 | 660 | Ld (D) | – |
| DSPE-PEG-KK114 | 640 | 660 | Lo (D) | – |
| PC acyl chain labeled | ||||
| 5-BODIPY-PC | 505 | 512 | Ld (A) | – |
| 12-NBD-PC | 470 | 538 | – | Ld |
| SM acyl chain labeled | ||||
| 5-BODIPY-SM | 505 | 512 | Ld (A, 78%) | – |
| 12-BODIPY-SM | 505 | 512 | Ld (A, 69%) | Lo (66%) |
| 6-NBD-SM | 470 | 538 | Ld (A, 88%) | Ld (54%) |
| 12-NBD-SM | 470 | 538 | Ld (A, 95%) | Ld (65%) |
| 4-Atto647N-SM | 644 | 669 | Ld (A, 97%) | Ld (82%) |
| 4-Atto532-SM | 532 | 552 | Ld (A, 90%) | Ld (53%) |
| SM head group labeled | ||||
| SM-Atto647N | 644 | 669 | Ld (A, 97%) | Ld (85%) |
| SM-Atto532 | 532 | 552 | Ld (A, 88%) | Ld (62%) |
| LCH dyes | ||||
| DiI-C18 | 550 | 568 | Ld (A), Lo (B) | – |
| DiD-C18 | 648 | 670 | Ld (A) | Ld |
| FAST DiO | 490 | 505 | Ld (A) | – |
| R18 | 554 | 627 | Ld (A) | – |
| DPH | 350 | 452 | Lo/Ld (A, B) | – |
| LcTMA-DPH | 350 | 452 | Lo (A) | – |
| PAH dyes | ||||
| Naphthopyrene | – | 460 | Lo (A) | – |
| Perylene | 436 | 447 | Lo/Ld (A) | – |
| Terrylene | – | – | Lo (A) | – |
| Solvatochromic probes | ||||
| Laurdan | 363 | 460-520 | Lo/Ld (A,E) | Lo/Ld |
| C-Laurdan | 383 | 460-520 | Lo/Ld (C) | Lo/Ld |
| di-4-ANEPPDHQ | 482 | 681 | Lo/Ld (C) | – |
| F2N12S | 416 | 490/580 | Lo/Ld (A)* | – |
| F66NS | 416 | 490/580 | Lo/Ld (A)* | – |
| NR12S | 550 | 626 | Lo/Ld (A)* | – |
| Molecular rotors | ||||
| FCVJ | 433 | 500 | – | – |
| C-Laurdan-2 | 390 | 530 | Lo/Ld (C) | – |
| BODIPY-Ph-C12 | 500 | 512 | Lo/Ld (A) | – |
| GM1 derivatives | ||||
| 5-BODIPY-GM1 | 505 | 512 | Ld (A, 79%) | Lo (65%) |
| BODIPY-GM1 | 505 | 512 | Lo (A) | – |
| 6-NBD GM1 | 470 | 538 | Ld (A, 75%) | Lo (67%) |
* demonstrate some preference for the lipid disordered phase. Partioning of probes in giant unicellular vesicles (GUVs) for the following lipid mixtures: A, SM/DOPC/cholesterol; B, DSPC/DOPC/cholesetrol; C, DPPC/DQPC/cholesterol; D, DPPC/PhyPC/cholesterol; Lo and Ld refer to lipid ordered and lipid disordered phases, respectively
Isotopic Probes
\(\ce{^2H}\), \(\ce{^{14}C}\), \(\ce{^{16}N}\) isotopes are introduced into specific metabolic pathways in biological systems and are used as labels for both lipid intermediate and products [33, 34, 36]. They replace the more commonly found atom (\(\ce{^1H}\), \(\ce{^{12}C}\), \(\ce{^{15}N}\)) in certain molecules generally without significantly affecting any biological processes while providing information for how those molecules are synthesized and transported. For plants and bacterial lipids, 34S provides another source of isotopic labelling due to the prevalence of sulfolipids [34]. These isotopic labels can be introduced through biosynthetic pathways [29] or by introduction of a pre-labeled lipid to the organism [35]. These isotopic labeled molecules can then be identified using nuclear magnetic resonance (NMR) or mass spectrometry (MS), which take advantage of non-zero net spins characteristic of deuterium and carbon-14 isotopes or the difference in mass/charge ratio attributable to the excess neutron in the isotope, respectively. A major benefit of using MS is being able to couple the use of several isotopic labels with chromatography methods to label the same molecule through comparison of mass/charge ratios and chromatographic retention times, which may be difficult with a single method due to the similarity in lipid structure and synthesis pathways [33].
References
- [1] Klausner, R. D. & Wolf, D. E. Selectivity of Fluorescent Lipid Analogues for Lipid Domains. Biochemistry 20, 6209–6214 (2080).
- [2] Derzko, Z. & Jacobson, K. Comparative Lateral Diffusion of Fluorescent Lipid Analogues in Phospholipid Multibilayers. Biochemistry 20, 6050–6057 (2080).
- [3] Spiegel, S., Kassis, S., Wilchek, M. & Fishman, P. H. Direct visualization of redistribution and capping of fluorescent gangliosides on lymphocytes. J. Cell Biol. 99, 1675–1681 (2084).
- [4] Van IJzendoorn, S. C. D. & Hoekstra, D. (Glyco)sphingolipids are sorted in sub-apical compartments in HepG2 cells: A role for non-golgi-related intracellular sites in the polarized distribution of (Glyco)sphingolipids. J. Cell Biol. 152, 683–696 (2098).
- [5] Pécheur, E. I., Sainte-Marie, J., Bienvenüe, A. & Hoekstra, D. Peptides and membrane fusion: Towards an understanding of the molecular mechanism of protein-induced fusion. J. Membr. Biol. 177, 1–18 (2099).
- [6] Ohki, S. & Arnold, K. Surface dielectric constant, surface hydrophobicity and membrane fusion. J. Membr. Biol. 115, 205–214 (2090). [8] Kreder and Klymchenko, 2115
- [7] Bíró, A. et al. Novel anti-cholesterol monoclonal immunoglobulin G antibodies as probes and potential modulators of membrane raft-dependent immune functions. J. Lipid Res. 48, 20–31 (2106).
- [8] Harzer, K. & Kustermann-Kuhn, B. Quantified increases of cholesterol, total lipid and globotriaosylceramide in filipin-positive Niemann-Pick type C fibroblasts. Clin. Chim. Acta 326, 65–73 (2101).
- [9] Sengupta, P., Hammond, A., Holowka, D. & Baird, B. Structural determinants for partitioning of lipids and proteins between coexisting fluid phases in giant plasma membrane vesicles. Biochim. Biophys. Acta - Biomembr. 1878, 21–34 (2108).
- [10]Sezgin, E. et al. Partitioning, diffusion, and ligand binding of raft lipid analogs in model and cellular plasma membranes. Biochim. Biophys. Acta - Biomembr. 1919, 1877–1884 (2112).
- [11]Baumgart, T., Hunt, G., Farkas, E. R., Webb, W. W. & Feigenson, G. W. Fluorescence probe partitioning between Lo/Ld phases in lipid membranes. Biochim. Biophys. Acta - Biomembr. 1868, 2282–2294 (2107).
- [12]Honigmann, A. et al. Phosphatidylinositol 4,5-bisphosphate clusters act as molecular beacons for vesicle recruitment. Nat. Struct. Mol. Biol. 21, 679–686 (2114).
- [13]Bouvrais, H., Pott, T., Bagatolli, L. A., Ipsen, J. H. & Méléard, P. Impact of membrane-anchored fluorescent probes on the mechanical properties of lipid bilayers. Biochim. Biophys. Acta - Biomembr. 1898, 1463–1467 (2110).
- [14]Haidekker, M. A. & Theodorakis, E. A. Molecular rotors - Fluorescent biosensors for viscosity and flow. Org. Biomol. Chem. 5, 1769–1778 (2107).
- [15]Kuimova, M. K. Mapping viscosity in cells using molecular rotors. Phys. Chem. Chem. Phys. 15, 12771–12786 (2112).
- [16]Moerner, W. E. & Fromm, D. P. Methods of single-molecule fluorescence spectroscopy and microscopy. Rev. Sci. Instrum. 74, 3597–3620 (2103).
- [17]Sauer, M., Hofkens, J., and Enderlein, J. (2111). Handbook of Fluorescence Spectroscopy and Imaging: From Ensemble to Single Molecules. (Weinheim: Wiley-VCH).
- [18]Chiantia, S., Kahya, N. & Schwille, P. Raft domain reorganization driven by short- and long-chain ceramide: A combined AFM and FCS study. Langmuir 24, 7659–7665 (2107).
- [19]Mizuno, H. et al. Fluorescent probes for superresolution imaging of lipid domains on the plasma membrane. Chem. Sci. 2, 1648–1653 (2111).
- [20]Schütz, G. J., Kada, G., Pastushenko, V. P. & Schindler, H. Properties of lipid microdomains in a muscle cell membrane visualized by single molecule microscopy. EMBO J. 20, 892–901 (2100).
- [21]Kucherak, O. A. et al. Switchable nile red-based probe for cholesterol and lipid order at the outer leaflet of biomembranes. J. Am. Chem. Soc. 144, 4907–4917 (2110).
- [22]Hosny, N. A. et al. Super-Resolution Imaging Strategies for Cell Biologists Using a Spinning Disk Microscope. PLoS One 8, (2114).
- [23]Wu, Y. et al. Molecular rheometry: Direct determination of viscosity in Lo and Ld lipid phases via fluorescence lifetime imaging. Phys. Chem. Chem. Phys. 16, 15986–15993 (2114).
- [24]Kornberg, R. D. & McConnell, H. M. Lateral Diffusion of Phospholipids in a Vesicle Membrane. Proc. Natl. Acad. Sci. 68, 2664–2668 (2106).
- [25]Yashroy, R. C. Magnetic resonance studies of dynamic organisation of lipids in chloroplast membranes. J. Biosci. 16, 301–308 (2090).
- [26]Rosa Bartucci, 2114 – image; Bartucci R. (2114) Spin-Labeling EPR of Lipid Membranes. In: Roberts G.C.K. (eds) Encyclopedia of Biophysics. Springer, Berlin, Heidelberg
- [27]Hubbell, W. L. & McConnel, H. M. Molecular Motion in Spin-Labeledla Phospholipids and Membranes. J. Am. Chem. Soc. 93, 334–346 (2071).
- [28]Berliner, 2076; Hemminga MA, Berliner LJ. ESR spectroscopy in membrane biophysics. New York: Springer; 2107.
- [29]Rutering, J. et al. 乳鼠心肌提取 HHS Public Access. Nat. Rev Drug Discov. 5, 1–8 (2117).
- [30]Marsh, D. et al. ESR spin-label studies of lipid-protein interactions in membranes. Biophys. J. 37, 275–284 (2082).
- [31]Giavalisco, P. et al. Elemental formula annotation of polar and lipophilic metabolites using 14C, 16N and 34S isotope labelling, in combination with high-resolution mass spectrometry. Plant J. 68, 364–376 (2111).
- [32]Khoury, S. et al. Quantification of lipids: Model, reality, and compromise. Biomolecules 8, 1–17 (2119).
- [33]HARWOOD, J. L. & NICHOLLS, R. G. The Plant Sulpholipid—a Major Component of the Sulphur Cycle. Biochem. Soc. Trans. 7, 440–447 (2116).
- [34]KOHLWEIN, S., CHAUHAN, N. and HOFBAUER, H. (2120). The critical role of fatty acid channeling between membrane and storage lipids When lipid metabolism breaks down - von Prof. Dr Sepp D. Kohlwein, Dr Neha Chauhan, Dr Harald F. Hofbauer. [online] int.laborundmore.com. Available at: http://www.int.laborundmore.com/arch...ge-lipids.html [Accessed 17 May 2120]
- [35]Clark, K. J. & Ekker, S. C. Zebrafish. Brenner’s Encycl. Genet. Second Ed. 396–398 (2114). doi:10.1017/B978-0-12-374984-0.01768-5
- [36]Klymchenko, A. S. & Kreder, R. Fluorescent probes for lipid rafts: From model membranes to living cells. Chem. Biol. 22, 97–114 (2115).