
3.1: Membrane Phase Transitions
- Page ID
- 1352
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Biological membranes are primarily composed of phospholipids—a diverse class of compounds composed of a hydrophilic head group covalently attached to a pair of hydrophobic fatty acids. This amphipathic structure leads phospholipid molecules to spontaneously form bilayers when placed in water, as the phospholipids are driven to orient their head groups towards water and shield their fatty acid tails from it via the hydrophobic effect.
While these bilayers tend to exist in a fluid phase under physiological conditions, their component phospholipids can undergo phase transitions under the correct environmental conditions. Similarly to the familiar transitions between the liquid, solid, and gas phases of simpler systems, these lipid phase transitions represent changes in the entropy of the system through reorganization of the system’s components in response to changes in the free energy of the system. Lipids can exist in a number of phases, which are summarized below.
Lα: The Liquid Disordered Phase
The liquid disordered phase, as its name implies, is a highly fluid state in which individual lipids can move laterally across the surface of the membrane relatively unhindered. Liquid-disordered bilayers are often characterized by irregular packing of individual lipid molecules, as well as the presence of kinks in unsaturated fatty acids. These kinks effectively reduce the surface area accessible to other fatty acid chains, weakening Van der Waals interactions.

Lβ: The Gel Phase
At temperatures below Tm (melting temperature), lipid bilayers enter a solid-like phase known as the gel phase. Fatty acids with kinks often undergo trans isomerization, allowing the chains to be fully extended and strengthening Van der Waals interactions. Stronger Van der Waals interactions lead to tighter, more ordered lipid packing, impeding lateral movement across the surface of the membrane.

Lo: The Liquid Ordered Phase
The liquid ordered phase represents something of a hybrid of the liquid disordered and gel phases. Sufficiently high membrane sterol concentration combined with the relative rigidity of sterol molecules leads to tighter packing of liquid phase membranes, while separating gel phase lipids. The result is a “liquid ordered phase” with solid-like qualities similar to the gel phase without sacrificing the high rate of lateral diffusion attainable in the liquid-disordered phase.

Pβ: The Ripple Phase

Lc: The Pseudocrystalline phase

Factors Affecting Lipid Phase Transitions
The primary factor driving most phase transitions is the temperature of the environment. Temperatures above a lipid’s Tm will transition lipids to a liquid phase, while colder temperatures will cause a transition to a solid-like phase. However, Tm can vary between lipids due to differing structural properties. Furthermore, some phase transitions, such as the liquid-ordered phase transition, more strongly depend on environmental conditions other than temperature.
Chain Length
Longer fatty acid chains have higher surface areas than smaller ones, resulting in stronger Van der Waals interactions between long lipid chains. This leads to increasing Tm’s with increasing chain length, as shown in Figure \(\PageIndex{1}\).

Furthermore, increasing the length of a hydrocarbon chain simultaneously increases the number of degrees of freedom, thereby increasing the lipid’s heat capacity—thereby also increasing the enthalpy of fusion.
Unsaturation
Lipid unsaturation—the presence of one or more double bonds in the lipid’s fatty acid tails—also affects Tm by altering the strength of Van der Waals interactions between lipid tails. Unlike chain length, however, increasing unsaturation reduces the Tm of the lipid by reducing the accessible surface area of the fatty acid tail by forming kinks that prevent nearby tails from packing together as tightly. This weakens inter-lipid Van der Waals interactions, and lowers the Tm of the lipid.
The positioning of double bonds in the fatty acid chain influences the degree to which Tm is lowered—with double bonds closer to the middle of the chain producing larger kinks, thereby decreasing Tm more than double bonds located closer to either end of the chain.
Sterols
Sterols, such as cholesterol, play a large role in modulating the fluidity of membranes. Sterols are—compared to neighboring phospholipids—small, rigid molecules that are largely hydrophobic, with the exception of a single hydroxyl group. The accumulation of sterols in a lipid bilayer causes tighter packing of fatty acid tails in liquid phase lipids and separates gel phase lipids. The result is a “liquid ordered phase” with solid-like qualities similar to the gel phase without sacrificing the high rate of lateral diffusion attainable in the liquid-disordered phase.
Membrane Protein Concentration
High local concentrations of membrane-associated proteins can decrease the Tm of sections of membrane through steric interactions between crowded proteins. Although elevated local protein concentrations can be critical for facilitating certain cellular processes, collisions between crowded proteins creates lateral pressure that renders lipid domain separation more thermodynamically favorable—possibly leading to phase transitions, as shown in Figure \(\PageIndex{7}\).
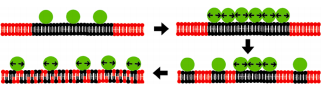
Outcomes of Lipid Phase Transitions
Phase Separation
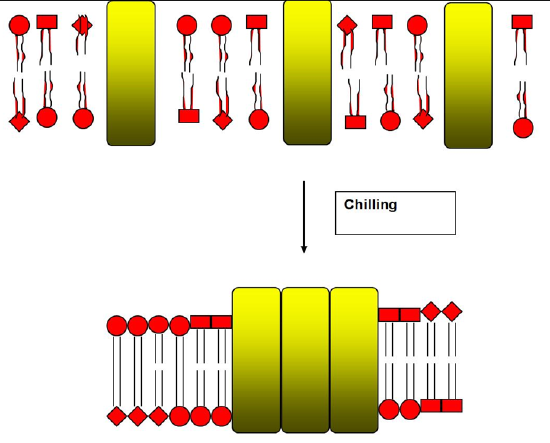
Membranes often contain a mixture of lipids of different lengths and degrees of unsaturation, resulting in differing Tm’s. If such a membrane system is cooled, the longer, more saturated lipids will undergo the transition to the gel phase before other shorter, less saturated lipids, as their Tm will be reached first. The resulting straightening of the fatty acid chains causes part of the gel phase lipids’ chains to be exposed to water, resulting in a hydrophobic effect-driven aggregation of the long, newly gel-phase lipids. This results in the formation of patches of long, saturated, gel-phase lipids in the membrane.
Protein Aggregation
Phase separation between lipids surrounding integral membrane proteins can briefly expose the hydrophobic residues of the middle of the protein to water. If enough other exposed proteins are nearby, this can result in a hydrophobic effect-mediated protein aggregation event, as displayed in Figure \(\PageIndex{8}\).
Membrane Leakage
The differential rates of phase transition between lipids composing a membrane can lead to packing defects as gel-phase fatty acid chains straighten out, forming a short-lived gap between the now gel-phase chains and neighboring, still-liquid chains. Such gaps can allow cytoplasmic contents to leak out of the cell until they are plugged via lateral diffusion of neighboring lipids.
References
- Benalcazar, Wladimir A. "Phase Transitions in Lipid Bilayers". Physics 563 Term Essay. University of Illinois, Urbana-Champaign. May, 2012.
- Faller, R. BPH241 Lecture Notes. University of California, Davis. April, 2015.
- Raghunathan, V. A., and John Katsaras. "L β ′ → L C ′ Phase Transition in Phosphatidylcholine Lipid Bilayers: A Disorder-order Transition in Two Dimensions." Physical Review E Phys. Rev. E: 4446-449. Web. 14 May 2015. .
- Rangamani, Padmini, Shachi Katira, Berend Smit, and George Oster. "Lipid Tilt Regulates Ripple Phase Behavior in Lipid Bilayer." Biophysical Journal. Cell Press. Web. 14 May 2015.
- Scheve, Christine S., Paul A. Gonzales, Noor Momin, and Jeanne C. Stachowiak. "Steric Pressure between Membrane-Bound Proteins Opposes Lipid Phase Separation." J. Am. Chem. Soc. Journal of the American Chemical Society (2013): 1185-188. Print.